
Gosh, it has been quite some time since I last posted an update.
Everyone I talk to on the MSc keeps saying how tired they are feeling, and I am no exception. I find it very hard to concentrate on getting work done, as I am distracted by the prospect of just finishing this course and getting out into the world, finding a decent job and (hopefully) earning a respectable wage for the first time in my life.
Still, I am struggling to stay grounded, to get the work done and my mind on the tasks in hand. I am about to throw myself into this research project. I have hundreds and hundreds of pages I need to read and understand in the coming weeks,and at the moment this task is quite daunting. Ho hum, it must be done.
Oh yeah, and we are just about to be set our final batch of coursework, and I should hopefully get my marks for that exam I had in January next week. Keep your fingers crossed for me!
Saturday, 27 February 2010
Monday, 15 February 2010
A proper post to follow, but in the mean time:
How might historical thinking about relationships between brain and language relate to current theories and interpretation of behavioural and neuroimaging data?
Introduction
There is no single canonical historical perspective on the relationship between brain and language; anecdotal reports of prototypical aphasic conditions were detailed by the ancient Egyptians, and many contradictory discoveries of dubious veracity emerged over the subsequent millennia. Therefore we shall consider the generally accepted model of the brain and language immediately prior to the twentieth century, shortly before the emergence of neuroscience as a formalised empirical pursuit, as the zenith of historical understanding of the subject, which we shall term the classical model. We then examine how this model relates to the predominant twentieth century assumptions of modularity and functional localisation, and in turn our current understanding of the brain. Finally, we consider what modern neuroscience has taught us that is beyond the technological capabilities of the historical pioneers of language.
The classical model of language
The two main contributors to the classical model of language were Paul Broca and Carl Wernicke, who pioneered our understanding of language production and comprehension respectively (Binder et al., 1997). Broca expanded on a hypothesis posited by his contemporary Jean-Baptiste Bouillaud to propose that the posterior inferior frontal gyrus of the left hemisphere was the region of the brain responsible for language production, based on lesion evidence gathered from his own patients. Wernicke identified that damage to the posterior part of the superior temporal gyrus in the left hemisphere resulted in a receptive aphasia with retained speech production, and postulated that this region was responsible for the memories of words and in turn the comprehension of the speech of others. These discoveries were historically significant in several respects; firstly they identified the left hemisphere as being the predominant hemisphere in language production and processing. Secondly, whilst these discoveries were a major step in the neurological understanding of language, they were even more significant for neuropsychology as a whole, as for the first time they elevated the concept of functional localisation above the pseudoscience of phrenology.
Whilst both Broca and Wernicke undoubtedly made a substantial contribution to our understanding of language, the classical model of language can be challenged in several respects. For example, lesions to the areas described by Broca and Wernicke do not always result in the deficits they described. Furthermore, aphasia of both production and comprehension of language can occur without lesions to the regions outlined in the classical model (Caplan, et al., 2007; Willmes, & Poeck, 1993). These discrepancies suggest that the neural basis of language is far more complex than can be accounted for by the historical model, meaning it must be either expanded or replaced. As a result language is still an intensely scrutinised field in modern neuroscience.
Language in the age of neuroscience
Early imaging studies somewhat supported the classical model of language (Cabeza & Nyberg, 2000), and more recently Broca’s area and the surrounding region have been shown to be active when translating phonemic information into articulatory information in preparation for speech (Papoutsi et al. 2009). The use of transcranial magnetic stimulation (TMS), when applied over Broca’s area in normal patients can temporarily prevent articulate speech, and almost paradoxically, the therapeutic use of TMS, when applied over the right hemispheric Broca’s area has been shown in some cases to improve picture naming in subjects with Broca’s aphasia (Naeser, et al., 2005). This behavioural data implicates Broca’s area in speech production, as the classical model dictates.
Broca was not without his critics, even in his own time, and shortly after Broca’s death the French neurologist Pierre Marie noted that expressive aphasia was not exclusively associated with lesions to Broca’s area, but also with regions such as the insula and the basal ganglia (Marie, 1906, as cited in Dronkers, Plaisant, Iba-Zizen, & Cabanis, 2007). Modern neuroscientists have also identified additional linguistic brain areas, including the thalamus, insula and basal ganglia (Damasio & Geschwind, 1984; Mazzocchi & Vignolo, 1979; Naeser & Helm-Estabrooks, 1985). This posthumous vindication for Marie’s remarkably prescient discovery is also somewhat ironic, as modern MRI studies have since shown significant damage to the insula and the basal ganglia in the preserved brains of Broca’s own patients (Dronkers et al., 2007).
The region commonly known as Wernicke’s area, the posterior superior temporal gyrus, has also been scrutinised by modern neuroscientists, and many have found evidence to support Wernicke’s hypothesis that this region plays a key role in language comprehension (Friederici, Makuuchi, & Bahlmann, 2009). An fMRI study by Grodzinsky and Friederici (2006) found this area to be involved in integration of syntactic and lexical information, crucial in decoding the speech of others. Increased activation of the posterior superior temporal gyrus has also been shown to be correlated with violations of grammatical structures, such as word order (Bornkessel, Zyssett, Friederici, von Cramon, Schlesewsky, 2005; Stowe, et al. 1998) and various other factors associated with the complexity of language comprehension tasks (Friederici, Makuuchi, & Bahlmann, 2009). This appears to support Wernicke’s hypothesis that this region is implicated in syntactic and lexical comprehension.
The modularity debate
The classical model of language was formed largely on the assumption of functional localisation. This theory suggested that specific cognitive faculties are functionally compartmentalised into distinct structures within the brain, and would later become entrenched in cognitive psychology by the likes of Fodor (1983), although many others subscribed to the concept of an integrated brain (Farah, 1994; Uttal, 2003). The modular doctrine has become somewhat challenged in light of advanced neuroimaging techniques, and some have speculated that the modular account of language must be drastically reconsidered, if not abandoned, in the light of modern imaging studies (Bates & Dick, 2000).
Contemporary neuroscientists have now identified a wide variety of structures within the brain that appear to be implemented in language production and processing (Fedorenko & Kanwisher, 2009). Furthermore, and perhaps uncomfortably for proponents of localisation, almost every part of the brain has at some point been prescribed a linguistic function by modern neuroscience (Bates & Dick, 2000). Whilst some of these studies support the classical model, many also flatly contradict it. For example, certain linguistic comprehension processes have been observed in regions independent of Wernicke’s area, including in Broca’s area (Ben-Shachar, Hendler, Kahn, Ben-Bashat, & Grodzinsky, 2003; Bornkessel-Schlesewsky, Schlesewsky, & von Cramon 2009) which would surely cause both of the behemoths of the classical model some distress. However, clear correlations have been observed between left hemispheric lesions and specific patterns of deficits (Cooper, Eichhorn, & Rodnitzky, 2008), therefore one might conclude that while it may be a gross oversimplification, there is almost certainly some specialisation for language in this region, and neither historical nor contemporary science has yet been able to fully explain how these interact with the brain as a whole.
The contributions of modern neuroscience
Contemporary scientific methods have helped to elucidate the linguistic brain in ways that the early pioneers, who could only examine the brain on post-mortem, were unable. Broca’s decision to avoid dissecting his subjects meant he grossly underestimated the extent of their subcortical damage, and the surface lesion had engulfed a far greater area of cortex than he had described (Dronkers et al., 2007), as they included additional parts of the frontal lobe and the lateral and medial prefrontal cortex, now known to also have important linguistic functions (Binder et al., 1997). This also explains why modern patients with lesions confined to Broca’s area often show rapid signs of recovery (Bakheit, Shaw, Carrington, & Griffiths, 2007), whilst Broca’s own patients symptoms continued to deteriorate until death. However, the recovery of modern aphasics also serves to illuminate a phenomenon which none of the historical researchers could have foreseen, neuroplasticity. Modern neuroimaging has revealed the ability of the brain to compensate for damage to many areas, including linguistic regions. For example, in patients with left hemispheric lesions the right hemisphere can often adopt some of the language functions normally associated with the left, and if the patient begins to recover from the aphasia the left hemisphere can begin to regain the lost language functions (Knopman, Rubens, Selnes, Klassen, & Meyer, 1984).
In infants, congenital damage to the left hemisphere does not typically result in aphasic conditions (Bates & Roe, 2001). Damage to the left hemisphere can lead to some developmental delays in language compared to damage to the right hemisphere but after infancy there is no significant difference in language deficit between those with congenital left or right damage (Bates, et al, 1997). Imaging studies have suggested that in cases such as these the right hemisphere can adapt to adopt the language faculties normally observed in the left hemisphere (Feldman, 2005). This strongly supports the assertion that whilst the left hemisphere is by default the predominant language centre of the brain, this is not a concrete configuration and can be developmentally subverted where necessary. Additionally, some linguistic functions have recently been identified in the right hemisphere in the normal population, such as the right homologue to Wernicke’s area, which is thought to process words with ambiguous meaning (Harpaz, Levkovitz, & Lavido, 2009). This has implications for the historical model of language, as it challenges the assertion that the left hemisphere is the categorical language centre of the brain, and in that it suggests that the functional areas described by Broca and Wernicke can be circumvented.
Modern neuroscience has also suggested that there is a strong association between Broca’s area and motor control (Jang, 2009), which is perhaps unsurprising given its close proximity to the primary motor regions of the brain. This may suggest that Broca’s aphasia may be at least partially a motor deficit rather than a strict language deficit. Although this seems to contradict the classical model, it was proposed by Broca’s contemporary, John Hughlings Jackson, as long ago as 1868 (Lorch, 2008). A motor association with Broca’s area may indicate that verbal language ability evolved as a result of motor expressions, such as hand gestures, that preceded it.
The limitations of comparing classical and contemporary models
The concept of language as defined by Broca, Wernicke and their contemporaries was arguably rather crude by the standards of modern neurolinguistics, as it consisted almost solely of expression and reception. The current interpretation of language is somewhat more nuanced, and encompasses concepts such as the difference between spoken and written language, semantic meaning, visual imagery, phonetics, working memory and non-verbal cues. For example, Fedorenko, Gibson, and Rohde (2006) examined the role of working memory in sentence comprehension, and Just (2008) observed that when we hear spoken sentences that prime mental imagery, activation occurs in the intraparietal sulcus. This level of scrutiny highlights the failure of the historical model to appropriately consider the complicated subcomponents of language. Therefore one could argue that historical and contemporary thinkers are not necessarily describing the same phenomena when they refer to language. In addition, the region referred to as Broca’s area varies considerably between studies and over time (Dronkers, et al., 2007), meaning many studies of language may not even be describing the same neural region as Broca discovered.
As a note of caution, both the techniques employed by classical and contemporary neuroscientists are to some extent methodologically flawed. One must use caution when using a lesioned brain to make inferences about normal functioning as we cannot guarantee that the unlesioned areas are functioning as normal (Rorden & Karnath, 2004), and we now know that many neural regions can adapt their function to compensate for damage elsewhere (Robertson & Murre, 1999), such as the right hemisphere adopting some of the lost language functioning resulting from lesions to the left hemisphere (Knopman, Rubens, Selnes, Klassen, & Meyer, 1984). Similarly, the interpretation of modern imaging techniques can be scientifically dubious. For example, the association between activation and cognitive processes can be ambiguous, it is unclear whether observed neural activity is related to excitation or inhibition of a particular function, and the arguments raised in the modularity debate may well confound any activity observed as most imaging studies assume a brain modular in function (Poldrack, 2006). For these reasons many imaging studies of language may well be methodologically unsound.
Conclusion
Although Broca and Wernicke introduced some of the key neural regions associated with language, these represent only a fraction of the brain regions that now seem to be involved in language, and the historical model was unable to speculate on the complex circuitry by which the many areas involved in production and comprehension interact. Neither Broca nor Wernicke was wrong per se, but the discoveries of each continue to be refined and re-evaluated in the light of technological and methodological advances. This is not to diminish their discoveries; the cumulative way in which science progresses means that the pioneering research conducted by the likes of Broca and Wernicke, and their unabashed willingness to disregard the received wisdom of the age, was a vital bedrock for the rapid acceleration in what we know today about language in the brain.
How might historical thinking about relationships between brain and language relate to current theories and interpretation of behavioural and neuroimaging data?
Introduction
There is no single canonical historical perspective on the relationship between brain and language; anecdotal reports of prototypical aphasic conditions were detailed by the ancient Egyptians, and many contradictory discoveries of dubious veracity emerged over the subsequent millennia. Therefore we shall consider the generally accepted model of the brain and language immediately prior to the twentieth century, shortly before the emergence of neuroscience as a formalised empirical pursuit, as the zenith of historical understanding of the subject, which we shall term the classical model. We then examine how this model relates to the predominant twentieth century assumptions of modularity and functional localisation, and in turn our current understanding of the brain. Finally, we consider what modern neuroscience has taught us that is beyond the technological capabilities of the historical pioneers of language.
The classical model of language
The two main contributors to the classical model of language were Paul Broca and Carl Wernicke, who pioneered our understanding of language production and comprehension respectively (Binder et al., 1997). Broca expanded on a hypothesis posited by his contemporary Jean-Baptiste Bouillaud to propose that the posterior inferior frontal gyrus of the left hemisphere was the region of the brain responsible for language production, based on lesion evidence gathered from his own patients. Wernicke identified that damage to the posterior part of the superior temporal gyrus in the left hemisphere resulted in a receptive aphasia with retained speech production, and postulated that this region was responsible for the memories of words and in turn the comprehension of the speech of others. These discoveries were historically significant in several respects; firstly they identified the left hemisphere as being the predominant hemisphere in language production and processing. Secondly, whilst these discoveries were a major step in the neurological understanding of language, they were even more significant for neuropsychology as a whole, as for the first time they elevated the concept of functional localisation above the pseudoscience of phrenology.
Whilst both Broca and Wernicke undoubtedly made a substantial contribution to our understanding of language, the classical model of language can be challenged in several respects. For example, lesions to the areas described by Broca and Wernicke do not always result in the deficits they described. Furthermore, aphasia of both production and comprehension of language can occur without lesions to the regions outlined in the classical model (Caplan, et al., 2007; Willmes, & Poeck, 1993). These discrepancies suggest that the neural basis of language is far more complex than can be accounted for by the historical model, meaning it must be either expanded or replaced. As a result language is still an intensely scrutinised field in modern neuroscience.
Language in the age of neuroscience
Early imaging studies somewhat supported the classical model of language (Cabeza & Nyberg, 2000), and more recently Broca’s area and the surrounding region have been shown to be active when translating phonemic information into articulatory information in preparation for speech (Papoutsi et al. 2009). The use of transcranial magnetic stimulation (TMS), when applied over Broca’s area in normal patients can temporarily prevent articulate speech, and almost paradoxically, the therapeutic use of TMS, when applied over the right hemispheric Broca’s area has been shown in some cases to improve picture naming in subjects with Broca’s aphasia (Naeser, et al., 2005). This behavioural data implicates Broca’s area in speech production, as the classical model dictates.
Broca was not without his critics, even in his own time, and shortly after Broca’s death the French neurologist Pierre Marie noted that expressive aphasia was not exclusively associated with lesions to Broca’s area, but also with regions such as the insula and the basal ganglia (Marie, 1906, as cited in Dronkers, Plaisant, Iba-Zizen, & Cabanis, 2007). Modern neuroscientists have also identified additional linguistic brain areas, including the thalamus, insula and basal ganglia (Damasio & Geschwind, 1984; Mazzocchi & Vignolo, 1979; Naeser & Helm-Estabrooks, 1985). This posthumous vindication for Marie’s remarkably prescient discovery is also somewhat ironic, as modern MRI studies have since shown significant damage to the insula and the basal ganglia in the preserved brains of Broca’s own patients (Dronkers et al., 2007).
The region commonly known as Wernicke’s area, the posterior superior temporal gyrus, has also been scrutinised by modern neuroscientists, and many have found evidence to support Wernicke’s hypothesis that this region plays a key role in language comprehension (Friederici, Makuuchi, & Bahlmann, 2009). An fMRI study by Grodzinsky and Friederici (2006) found this area to be involved in integration of syntactic and lexical information, crucial in decoding the speech of others. Increased activation of the posterior superior temporal gyrus has also been shown to be correlated with violations of grammatical structures, such as word order (Bornkessel, Zyssett, Friederici, von Cramon, Schlesewsky, 2005; Stowe, et al. 1998) and various other factors associated with the complexity of language comprehension tasks (Friederici, Makuuchi, & Bahlmann, 2009). This appears to support Wernicke’s hypothesis that this region is implicated in syntactic and lexical comprehension.
The modularity debate
The classical model of language was formed largely on the assumption of functional localisation. This theory suggested that specific cognitive faculties are functionally compartmentalised into distinct structures within the brain, and would later become entrenched in cognitive psychology by the likes of Fodor (1983), although many others subscribed to the concept of an integrated brain (Farah, 1994; Uttal, 2003). The modular doctrine has become somewhat challenged in light of advanced neuroimaging techniques, and some have speculated that the modular account of language must be drastically reconsidered, if not abandoned, in the light of modern imaging studies (Bates & Dick, 2000).
Contemporary neuroscientists have now identified a wide variety of structures within the brain that appear to be implemented in language production and processing (Fedorenko & Kanwisher, 2009). Furthermore, and perhaps uncomfortably for proponents of localisation, almost every part of the brain has at some point been prescribed a linguistic function by modern neuroscience (Bates & Dick, 2000). Whilst some of these studies support the classical model, many also flatly contradict it. For example, certain linguistic comprehension processes have been observed in regions independent of Wernicke’s area, including in Broca’s area (Ben-Shachar, Hendler, Kahn, Ben-Bashat, & Grodzinsky, 2003; Bornkessel-Schlesewsky, Schlesewsky, & von Cramon 2009) which would surely cause both of the behemoths of the classical model some distress. However, clear correlations have been observed between left hemispheric lesions and specific patterns of deficits (Cooper, Eichhorn, & Rodnitzky, 2008), therefore one might conclude that while it may be a gross oversimplification, there is almost certainly some specialisation for language in this region, and neither historical nor contemporary science has yet been able to fully explain how these interact with the brain as a whole.
The contributions of modern neuroscience
Contemporary scientific methods have helped to elucidate the linguistic brain in ways that the early pioneers, who could only examine the brain on post-mortem, were unable. Broca’s decision to avoid dissecting his subjects meant he grossly underestimated the extent of their subcortical damage, and the surface lesion had engulfed a far greater area of cortex than he had described (Dronkers et al., 2007), as they included additional parts of the frontal lobe and the lateral and medial prefrontal cortex, now known to also have important linguistic functions (Binder et al., 1997). This also explains why modern patients with lesions confined to Broca’s area often show rapid signs of recovery (Bakheit, Shaw, Carrington, & Griffiths, 2007), whilst Broca’s own patients symptoms continued to deteriorate until death. However, the recovery of modern aphasics also serves to illuminate a phenomenon which none of the historical researchers could have foreseen, neuroplasticity. Modern neuroimaging has revealed the ability of the brain to compensate for damage to many areas, including linguistic regions. For example, in patients with left hemispheric lesions the right hemisphere can often adopt some of the language functions normally associated with the left, and if the patient begins to recover from the aphasia the left hemisphere can begin to regain the lost language functions (Knopman, Rubens, Selnes, Klassen, & Meyer, 1984).
In infants, congenital damage to the left hemisphere does not typically result in aphasic conditions (Bates & Roe, 2001). Damage to the left hemisphere can lead to some developmental delays in language compared to damage to the right hemisphere but after infancy there is no significant difference in language deficit between those with congenital left or right damage (Bates, et al, 1997). Imaging studies have suggested that in cases such as these the right hemisphere can adapt to adopt the language faculties normally observed in the left hemisphere (Feldman, 2005). This strongly supports the assertion that whilst the left hemisphere is by default the predominant language centre of the brain, this is not a concrete configuration and can be developmentally subverted where necessary. Additionally, some linguistic functions have recently been identified in the right hemisphere in the normal population, such as the right homologue to Wernicke’s area, which is thought to process words with ambiguous meaning (Harpaz, Levkovitz, & Lavido, 2009). This has implications for the historical model of language, as it challenges the assertion that the left hemisphere is the categorical language centre of the brain, and in that it suggests that the functional areas described by Broca and Wernicke can be circumvented.
Modern neuroscience has also suggested that there is a strong association between Broca’s area and motor control (Jang, 2009), which is perhaps unsurprising given its close proximity to the primary motor regions of the brain. This may suggest that Broca’s aphasia may be at least partially a motor deficit rather than a strict language deficit. Although this seems to contradict the classical model, it was proposed by Broca’s contemporary, John Hughlings Jackson, as long ago as 1868 (Lorch, 2008). A motor association with Broca’s area may indicate that verbal language ability evolved as a result of motor expressions, such as hand gestures, that preceded it.
The limitations of comparing classical and contemporary models
The concept of language as defined by Broca, Wernicke and their contemporaries was arguably rather crude by the standards of modern neurolinguistics, as it consisted almost solely of expression and reception. The current interpretation of language is somewhat more nuanced, and encompasses concepts such as the difference between spoken and written language, semantic meaning, visual imagery, phonetics, working memory and non-verbal cues. For example, Fedorenko, Gibson, and Rohde (2006) examined the role of working memory in sentence comprehension, and Just (2008) observed that when we hear spoken sentences that prime mental imagery, activation occurs in the intraparietal sulcus. This level of scrutiny highlights the failure of the historical model to appropriately consider the complicated subcomponents of language. Therefore one could argue that historical and contemporary thinkers are not necessarily describing the same phenomena when they refer to language. In addition, the region referred to as Broca’s area varies considerably between studies and over time (Dronkers, et al., 2007), meaning many studies of language may not even be describing the same neural region as Broca discovered.
As a note of caution, both the techniques employed by classical and contemporary neuroscientists are to some extent methodologically flawed. One must use caution when using a lesioned brain to make inferences about normal functioning as we cannot guarantee that the unlesioned areas are functioning as normal (Rorden & Karnath, 2004), and we now know that many neural regions can adapt their function to compensate for damage elsewhere (Robertson & Murre, 1999), such as the right hemisphere adopting some of the lost language functioning resulting from lesions to the left hemisphere (Knopman, Rubens, Selnes, Klassen, & Meyer, 1984). Similarly, the interpretation of modern imaging techniques can be scientifically dubious. For example, the association between activation and cognitive processes can be ambiguous, it is unclear whether observed neural activity is related to excitation or inhibition of a particular function, and the arguments raised in the modularity debate may well confound any activity observed as most imaging studies assume a brain modular in function (Poldrack, 2006). For these reasons many imaging studies of language may well be methodologically unsound.
Conclusion
Although Broca and Wernicke introduced some of the key neural regions associated with language, these represent only a fraction of the brain regions that now seem to be involved in language, and the historical model was unable to speculate on the complex circuitry by which the many areas involved in production and comprehension interact. Neither Broca nor Wernicke was wrong per se, but the discoveries of each continue to be refined and re-evaluated in the light of technological and methodological advances. This is not to diminish their discoveries; the cumulative way in which science progresses means that the pioneering research conducted by the likes of Broca and Wernicke, and their unabashed willingness to disregard the received wisdom of the age, was a vital bedrock for the rapid acceleration in what we know today about language in the brain.
Thursday, 11 February 2010
And relax...
I just submitted 2 major pieces of coursework I have been working on for some time, what a relief.
Over the last week I have become somewhat obsessed. On Monday, I took part in a neuroimaging experiment which involved going in an MRI machine. The lady who conducted the study was nice enough to let me keep the data file produced by my scan.
Which means I can now do all sorts of fancy things on my laptop, not just make images like this:

But also make fun videos like this:
And, I just discovered, I can convert the scan in 3D, which looks a bit like this:
Look out for the upcoming guided tour to my brain.
What, you think I'm joking?
Over the last week I have become somewhat obsessed. On Monday, I took part in a neuroimaging experiment which involved going in an MRI machine. The lady who conducted the study was nice enough to let me keep the data file produced by my scan.
Which means I can now do all sorts of fancy things on my laptop, not just make images like this:
But also make fun videos like this:
And, I just discovered, I can convert the scan in 3D, which looks a bit like this:
Look out for the upcoming guided tour to my brain.
What, you think I'm joking?
Monday, 1 February 2010
On Sunday I got my first glimpse of some serious neuroimaging machinery.
My supervisor invited me along to watch him testing a participant in an experiment that is very similar to the one he and I will be conducting in a few months time.
The participant was required to under go an MEG, or magnetoencephalogram.
Just like this one...
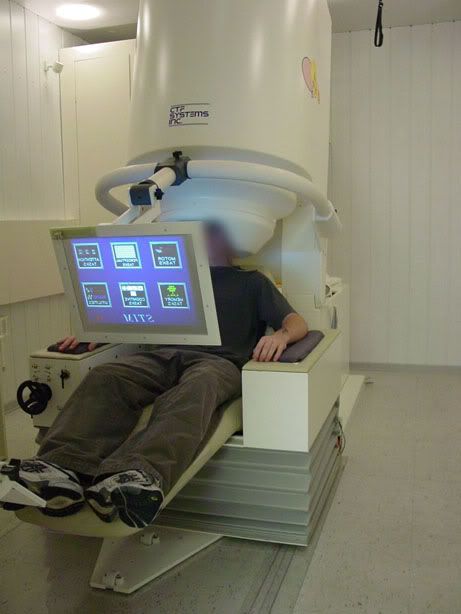
Although MEG is not strictly an imaging tool, it does allow us to view brain waves in different parts of the cerebral cortex. It works by recording the electrical activity at over 270 points in the brain. Once this is done you can examine the data to see what activity correlated with the task the participant was doing. Clever stuff.
The participant was sat in the machine and played a series of tones, at 2 different pitches, some of which were doubled in length. He had to press a button every time he heard a longer tone.
Halfway through the test one of the tones became associated with an electric shock to the forearm.
Christian didn't explain to me the exact nature of this study, but it is very similar in nature to the one we proposed (a few entries back in this very blog, if you want to read about it).
Oh, and I've volunteered to take part myself next week, and in addition I might be taking part in an fMRI experiment too, which I will tell you all about if it comes to pass.
Exciting times!
My supervisor invited me along to watch him testing a participant in an experiment that is very similar to the one he and I will be conducting in a few months time.
The participant was required to under go an MEG, or magnetoencephalogram.
Just like this one...
Although MEG is not strictly an imaging tool, it does allow us to view brain waves in different parts of the cerebral cortex. It works by recording the electrical activity at over 270 points in the brain. Once this is done you can examine the data to see what activity correlated with the task the participant was doing. Clever stuff.
The participant was sat in the machine and played a series of tones, at 2 different pitches, some of which were doubled in length. He had to press a button every time he heard a longer tone.
Halfway through the test one of the tones became associated with an electric shock to the forearm.
Christian didn't explain to me the exact nature of this study, but it is very similar in nature to the one we proposed (a few entries back in this very blog, if you want to read about it).
Oh, and I've volunteered to take part myself next week, and in addition I might be taking part in an fMRI experiment too, which I will tell you all about if it comes to pass.
Exciting times!
Subscribe to:
Comments (Atom)